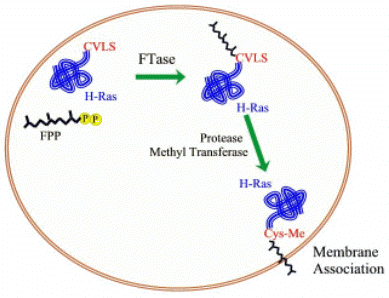
|
diphosphate onto protein substrates in
the cell (Click here for FTase chemical reaction).
Farnesylation is a form of post-translational protein lipidation,
which also includes geranylgeranylation, myristoylation, and acylation.
Addition of a lipid to the protein increases its hydrophobicity, which
increases the affinity of the protein substrate for the cell membrane, or
possibly a membrane associated receptor. Therefore, these modifications
play an important role in cell signaling. Known substrates of FTase include
H-Ras, K-Ras, N-Ras, RhoB, CENP-E and CENP-F. These proteins are all
involved in cell proliferation, and it has been shown that inhibiting Ras
farnesylation renders Ras inactive in the cytosol. Mutated Ras is found in
30% of human cancers, trapped in its active (GTP-bound) conformation so it
signals for hyperproliferation of cells to form tumors. The short
circuiting of the Ras pathway by farnesyltransferase inhibitors (FTIs) has
been a tremendous area of research in the past ten years, creating over 300
patents and three compounds currently in clinical trials.
FTase is part of a unique class of enzymes that
catalyze zinc-mediated sulfur alkylation. This class includes methionine
synthase (cobalamin dependent and independent), methyltransferases, the E.
coli DNA repair enzyme Ada,
and geranylgeranyltransferase I and II. It is not yet well understood how
these enzymes catalyze this type of reaction. These enzymes have
surprisingly different zinc coordination spheres considering their similar
reactions, ranging from four cysteines (Ada)
to only one (FTase, GGTase). The prenyltransferases (FTase, GGTase) share a
unique Asp-Cys-His-H2O zinc site. The water is displaced when
the cysteine sulfur of the protein substrate coordinates to the zinc (1).
This metal coordination lowers the pKa of the sulfur so that it
is deprotonated at physiological pH, enhancing the reactivity of the
substrate (2). This zinc does not only react with proteins, but also
several small thiol-containing compounds in vitro (3). In addition to this catalytic zinc, the
mechanism of FTase is also dependent on a magnesium ion. This metal is
thought to bind to the pyrophosphate of the substrate farnesyl diphosphate
(FPP), stabilizing it as a leaving group by neutralizing the negative
charge (4). This increases the rate of chemistry 700-fold. The reaction
proceeds through an exploded transition state, with both nucleophilic and
carbocation character (5).
PubMed
Search (most recent publications)
Collaborators on this project
Patrick Casey, Duke
University, NC
Matthew Saderholm,
Berea College, KY
Richard A Gibbs,
Purdue University, IN
James E
Penner-Hahn, University of Michigan, MI
Dale
Poulter, University of Utah, UT
|