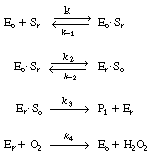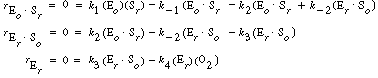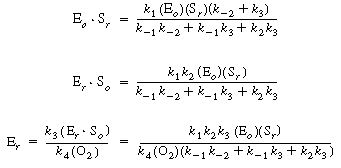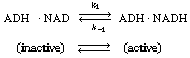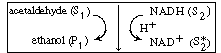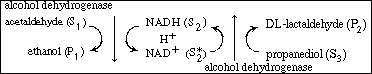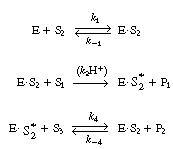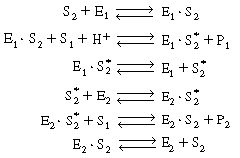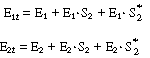Chapter 9: Reaction Mechanisms, Pathways, Bioreactions and Bioreactors
Professional Reference Shelf
The following information is taken from the 4th edition of Elements of Chemical Reaction Engineering, so the equation numbers correspond to those found in that book
R9.6.1: Microbial Growth on Multiple Substrates |
||||
R9.6.2 Enzyme Regeneration |
||||
|
As a first example we shall consider the oxidation of glucose (Sr
) with the aid of the enzyme glucose oxidase [represented as either G.O.
or (Eo )] to give
|
||||
|
In this reaction, the reduced form of glucose oxidase (G.O.H2 ), which will be represented by Er , cannot catalyze further reactions until it is oxidized back to Eo . This oxidation is usually carried out by adding molecular oxygen to the system so that glucose oxidase, Eo , is regenerated. Hydrogen peroxide is also produced in this oxidation regeneration step. |
||||
|
|
||||
| Overall, the reaction is written | ||||
|
|
||||
|
In biochemistry texts, reactions of this type involving regeneration are usually written in the form |
||||
|
|
||||
| The reaction is believed to proceed by the following sequence of elementary reactions: | ||||
|
|
||||
| We shall assume that reaction involving the dissociation
between reduced glucose oxidase and |
||||
|
|
(R7.4-1) | |||
| After applying the pseudo-steady-state hypothesis to the rates of
formation of (Eo |
||||
|
|
||||
| we can solve for the following concentrations of the active intermediates in terms of the concentrations of glucose, oxygen, and unbound oxidized enzyme. | ||||
|
|
(R7.4-2) (R7.4-3) (R7.4-4) |
|||
| After substituting Equation (R7.4-3) into Equation (R7.4-1), the rate law is written as | ||||
|
|
(R7.4-5) | |||
| The total enzyme initially present is given by the sum | ||||
|
(Et ) = (Eo
) + (Eo |
(R7.4-6) | |||
| After using Equations (R7.4-2), (R7.4-3), and (R7.4-4) to substitute for the active intermediate in Equation (R7.4-6), one can then solve for the unbound oxidized enzyme concentration, Eo , and substitute it into Equation (R7.4-5) to obtain the form of the rate law | ||||
|
|
(R7.4-7) | |||
|
Example R9.6-1 |
||||
| The reaction above illustrates how an enzyme can be regenerated through the addition of another substrate, in this case O2 . | ||||
R9.6.3 Enzyme Cofactors |
||||
| In many enzymatic reactions, and in particular biological reactions, a second substrate (i.e., species) must be introduced to activate the enzyme. This substrate, which is referred to as a cofactor or coenzyme even though it is not an enzyme as such, attaches to the enzyme and is most often either reduced or oxidized during the course of the reaction. The enzyme-cofactor complex is referred to as a holoenzyme. An example of the type of system in which a cofactor is used is the formation of ethanol from acetaldehyde in the presence of the enzyme alcohol dehydrogenase (ADH) and the cofactor nicotinamide adenine dinucleotide (NAD). After the enzyme is activated by combination with the cofactor in its reduced state, NADH, | ||||
|
|
||||
| the holoenzyme (ADH (ADH |
||||
|
|
||||
| The inactive form of the enzyme-cofactor complex for a specific
reaction and reaction direction is called an apoenzyme. This reaction
is followed by dissociation of the apoenzyme (ADH |
||||
|
|
||||
| The values of the specific rates are3 : | ||||
|
|
||||
|
Typical initial concentrations for a small laboratory batch reactor
experiment might be alcohol dehydrogenase |
||||
|
|
||||
| The ADH enzyme molecule produced by the dissociation of (ADH |
||||
|
|
||||
| rather than having the enzyme ADH go through the steps of dissociating from | ||||
|
|
||||
| and then combining with NADH: | ||||
|
|
||||
|
Example R9.6-2 |
||||
| Equation (RE7.4-1) is of a form that is often used in the interpretation of initial rate data for enzymatic reactions involving two substrates. The parameters K12 , K1 , K2 , and Vmax in Equation (RE7.4-1), which was first developed by Dalziel,4 may be evaluated through a series of Lineweaver-Burk plots. After substituting the numerical values for K1 , K2 , K12 and recalling the initial concentrations specified (S1,0 = 0.1 g mol/L, S2,0 = 10-4 g mol/L), we see that we can neglect K12 and K2 (S2 ) with respect to the other terms in the denominator, in which case Equation (G25-7) becomes | ||||
|
|
(R7.4-8) | |||
| The initial rate is rp 0
= 3.7 x 10-6 g mol/L |
||||
R9.6.4 Multiple-Substrate Systems In the preceding section we stated that the rate of formation of ethanol
might be increased if the (ADH |
||||
|
|
||||
| As a first approximation, this sequence of reactions could be represented by the following elementary steps: | ||||
|
|
||||
|
Example R9.6-3 This same reaction has been carried in the reverse direction
by Gupta and Robinson.5
They measured the initial rate of conversion of DL-lactaldehyde to propanediol
in the presence of NAD 1 and ADH. The
rate of dissociation of the enzyme-cofactor complex (ADH
|
||||
| In analyzing multiple reactions in this manner, one should always
question the validity of the application of the PSSH to the various active intermediates.
Since the nicotinamide adenine dinucleotide is continually regenerated and the total concentration of the cofactor (in its oxidized, reduced, bound, and unbound forms) remains constant throughout the course of the reaction, it might be desirable to replace S2 in the rate law in terms of the total cofactor concentration, S t . Neglecting any unbound, the total (initial) cofactor concentration is |
||||
|
|
(R7.4-9) | |||
| The total concentration of enzyme is | ||||
| nbsp; |
|
(R7.4-10) | ||
| Subtracting Equation (R7.4-9) from Equation (R7.4-10), we obtain | ||||
|
|
(R7.4-11) | |||
| Equation (R7.4-11) can be rewritten in the form | ||||
|
|
(R7.4-11) | |||
| where | ||||
|
|
(R7.4-12) | |||
| After adding Equations (R7.4-11) and (R7.4-12) and rearranging, we obtain | ||||
|
|
(R7.4-13) | |||
| which is solved for the unbound enzyme concentration in terms of S1 , S3 , P2 , Et , and St . | ||||
|
|
(R7.4-14) | |||
| We can substitute Equation (R7.4-14) into (R7.4-9) and rearrange to determine the concentration of the unbound cofactor in its reduced form, that is, | ||||
|
|
(R7.4-15) | |||
| The rate law was given by | ||||
|
|
(R7.4-16) | |||
| One could substitute Equations (R7.4-14) and (R7.4-15) for S2 and E into Equation (R7.4-16) to arrive at a reasonably complicated rate law involving S1 , St , S 3 , P2 , and Et . However, a computer solution would be used in most reaction sequences that are this involved algebraically, in which case further substitution would not be necessary and one could use Equations (R7.4-14), (R7.4-15), and (R7.4-16) directly. | ||||
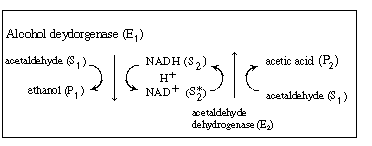
R9.6.5 Multiple Enzymes SystemsWe shall again consider the production of ethanol from acetaldehyde which uses the cofactor NADH. However, the regeneration of NAD+ to NADH is brought about in a reaction catalyzed by acetaldehyde dehydrogenase (E2 ), which produces acetic acid from acetaldehyde: |
||||
|
|
||||
|
This sequence can be written in abstract notation as |
||||
|
|
||||
| We could apply the PSSH to E1 |
||||
|
|
||||
| in deriving the rate law for this system. We shall not carry through
the necessary algebraic manipulation to obtain the rate law here, as all the principles
for determining the rate law have been presented, and there would be little more to
be accomplished by doing this. The sections on enzymatic reactions were meant to serve as a brief, yet somewhat encompassing discussion of enzyme kinetics. Further discussion can be found in the Supplementary Reading for Chapter 9. |